TAXONOMY
The taxonomy of Quercus genus is extremely complex as a result of high species numbers, wide geographical distribution, great morphological variability, as well as widespread hybridization between infrageneric taxa and changes in morphological features. There is so much variation within species that even the concept of species has been questioned by several authors (Burger, 1975; Van Valen, 1976) and further complications in taxonomy are due to frequent interspecific hybridization. As a result, the classification of oaks has been historically a matter of debate, from ∅rsted to Nixon more than 20 classifications have been proposed. Up to date, the most taxonomically valuable morphological features of Quercus species were foliar and fruit characteristics; hence, the taxonomic classifications of oaks were usually based only on these features (Wrońska-Pilarek et al., 2016)
Because of the immense size and wide distribution of oaks, authors did not always agree, about the classification or about the criteria to be used for it. In addition, classifications are often limited to a restricted part of the distribution of the genus that constitutes an additional heterogeneity factor (Denk et al., 2017a). One of the most complete classifications, as much for its geographic as specific cover, is that of Camus (1936-1954), her classification criteria were mostly based on foliar and fruit characteristics. In Camus’ classification the genus Quercus (sensu lato) is subdivided in two sub genera: sub genus Euquercus (Quercus sensu stricto) and sub genus Cyclobalanopsis. There are about 150 species belonging to Cyclobalanopsis which exist only in South Asia, whereas species belonging to Euquercus are the more familiar oak species. The sub genus Euquercus (Quercus sensu stricto) was further subdivided into 6 different sections by Camus (1936-1954).
Earlier taxonomists have suggested that the genus Quercus is heterogeneous enough that it should be divided into two or more genera. Schottky (1912), Trelease (1924) Hjelmqvist (1948), Brett (1964), Liu and Liao (1976). Schwarz (1936, 1964) even divided Quercus into four separate genera. Others, in contrast, have suggested that Quercus should be united to the genus Lithocarpus (De Candolle, 1868; Bentham & Hooker, 1880; Corner, 1939).
In comparison to Camus’ classification, the section Cerris (sensu Camus) was divided into additional sections by Asian botanists (section Brachylepides and Aegilops by Zheng, 1985, for example). Zheng also introduced a new section Englerianae merging species that belonged to sections Lepidobalanus and Cerris (sensu Camus). In the same way, Schwarz (1964) equally proposed to individualize the subsection Cocciferae of section Cerris (sensu Camus) with the specie Q. ilex into section Sclerophyllodrys.
The access to DNA markers and sequences provided new tools to resolve the taxonomical classification based on phylogenetic approaches. Different studies applying chloroplast DNA (cpDNA) and low-copy nuclear gene (LCNG) data have recovered a provisional framework phylogeny for the genus Quercus and identified the relative position of the genus within the Fagaceae (Manos et al., 1999 and Kremer et al., 2012). These studies succeed in identifying a predominantly American clade within the genus Quercus, containing sections Quercus (the white oaks, including the Virentes from the Americas and the Roburoids from Eurasia), Lobatae (the red), and the Protobalanus (the intermediate oaks/golden oaks). The fossil records and additional molecular analyses using rDNA-ITS and 5S-IGS sequences established the divergence of sections Quercus and Lobatae at a minimum of 23–33 Mya and define this clade excluding the Cerris and Ilex clades of Eurasia (Hipp et al., 2014).
More recently two nuclear-encoded molecular markers, the widely used ITS region of the 35S rDNA cistron and a 2415 bp long part of the CRABS CLAW gene (CRC), have clarified the infrageneric classification by recognizing five or six major groups (Manos et al., 2001; Oh & Manos, 2008; Denk & Grimm, 2010) that are mostly in agreement with the classification systems of Camus (1936-1954), Schwarz (1936) and Menitsky (1984). The North American group Protobalanus, the group Lobatae, the group Quercus, the Eurasian Group Cerris and Ilex (according to Denk & Grimm, 2010) and the East Asian group Cyclobalanopsis. These six infrageneric groups were later confirmed with a larger set of nuclear encoded genes (Hubert et al., 2014). These infrageneric groups are also recognized using pollen morphology, particularly based on their ornamentation (; Denk et al., 2017b). Previous Subgeneric classifications based on morphological characters are potentially at odds with this new pattern, but the genus Quercus in the broad sense still remains united by floral synapomorphies and a circular single fruit or acorn surrounded by conspicuous accessories.

Quercus sessile [ = Quercus petraea]. Photo by Antoine Kremer.
The New Establish Classification
Conventionally, until very recently, the central/eastern European tradition followed in principle the classification system of Schwarz (1936), whereas the western/southern European tradition relied on the monographic work of Camus (1936-1954).
The molecular phylogeny proposed by Manos et al.’s (2001) challenges the traditional views from Ørsted until Nixon. According to this, the intermediate and white oaks grouped with the red oaks, forming the “New World Clade”, but not with the Cerris and Ilex oaks. The latter formed an “Old World Clade” that later would be shown to include the cycle-cup oaks. While the red oaks and cycle-cup oaks were resolved in well-supported and distinct clades within their respective subtrees, the situation appeared more complex for Camus section Cerris (including a few Ilex oaks) and the white oaks.
One of the most profound recent changes, consequence of the advent of molecular phylogenetics, has been the realization that the traditional split between subgenus Cyclobalanopsis (East Asian) and subgenus Quercus (all other oaks) is artificial (Manos et al., 2008). At present, the overall signal is in line with the “New World/Old World Clade” concept introduced by Oh and Manos (2008) that is the groups: subgenus Quercus and subgenus Cerris.
The most recent classification is by Denk et al. (2017a) based on diverse works for diagnostic morphological characters for the recognized groups of oaks; from pollen morphology, updated information on the position of aborted ovules, DNA sequence data (nuclear encoded genes) and from a nuclear reduced representation of next-generation sequencing (RADseq) data set (Oh and Manos, 2008; Hubert et al., 2014; Hipp et al. 2015).
Classification by Denk and collaborators:


Comparison of the newest classification proposed by Denk and collaborators with the most important ones before it:
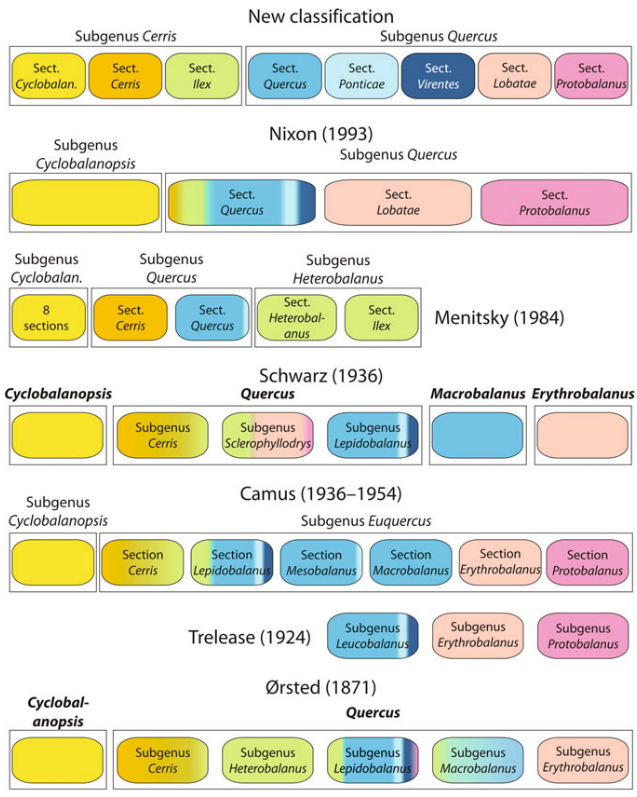
*Color gradients are proportional, reflecting the proportion of species with different systematic affiliation according to Denk et al. (2017a) included in each taxon. Names in bold are treated as genera. Schemes taken from Denk et al. (2017a).